Артемизинин е лекарствена билка, която се използва широко за лечение на малария. Последните проучвания показват, че тази „чудотворна билка“ има противоракови ефекти и може също да стимулира имунната система!
Въведение
Артемизинин (ART) е растително производно съединение, изолирано от Artemisia annua, сладък пелин, билка използвана в китайската билкова медицина.
Това съединение (заедно с производните му лекарства), е препоръчано от Световната здравна организация при лечението на малария, причинена от Plasmodium фалципарум ( R ).
Лекарството е било толкова ефективно при лечението на болестта, че откриващият Youyou Tu е получил Нобеловата награда за физиология или медицина ( R ).
Редица полусинтетични производни се получават от тази билка за използване при борба програми малария, най-добре между тях включват Артеметер, Артезунат и Артенимол (β-дихидро-артемизинин, DHA) ( R ).
Ползи за здравето
1) Артемизинин е ефективно анти-маларийно вещество и предпазва от други инфекции
Артемизинин е около 90% ефективно при лечението на неусложнена малария, причинена от Plasmodium фалципарум ( R ). Въпреки това широкото използване на лекарствената монотерапия води до развитие на резистентност при маларийни паразити.
Когато това лекарство се използва в комбинация с други антималарийни лекарства, P. falciparum е много малко вероятно да стане резистентен към него, поради което СЗО препоръча употребата на комбинация от Артемизинин (ACT) ( R ).
Екстрактите на Артемизининът имат антимикробна активност срещу пародонтопатични микроби като Aggregatibacter actinomycetemcomitans , Fusobacterium nucleatum subsp. animalis , Fusobacterium nucleatumsubsp. polymorphum и Prevotella посредничество ( R ).
Той намалява процентите на репликация на вирусите на хепатит В и С ( R ) и редица човешки херпес вирус ( R1 , R2 , R3 ).
2) Artemisinin помага за предотвратяване на рака
В едно рандомизирано проучване Артесунат контролирано комбинира с NP (режим на химиотерапия на винорелбин и цисплатин) може да повиши скоростта на краткосрочна оцеляване и удължено ТРР (време до прогресия) на пациенти с напреднал недребноклетъчен белодробен рак (NSCLC) ( R , R2 ).
Дългосрочното лечение на двама пациенти с метастатичен улеален меланом с Артемизинин в комбинация със стандартна химиотерапия показва значително подобрение ( R ).
Лечението с Артеметер е от полза за подобряване на качеството на живот на 75-годишен мъжки пациент с хипофизни макроаденоми ( R ).
Артемизинин и Resveratrol притежават синергичен антитуморен ефект, тази комбинация може да бъде ефективна терапевтична стратегия за рак ( R ).
ART инхибира растежа на клетъчните линии на жлъчния мехур (чрез индуциране на ROS и блокиране на клетъчния цикъл), поради което може да бъде подходящ за лечение на рак на жлъчния мехур ( R1 , R2 ).
Той индуцира антипролиферативни и проапоптотични ефекти при инфектирани с HPV-39 човешки тумори на маточната шийка ( R ).
Новите производни на ART (AD1-AD8) имат висока антипролиферативна активност срещу ракови клетки на черния дроб / колона , е необходима по-нататъшна оценка при животински модели ( R1 , R2 ).
3) Артемизининът е противовъзпалителен и имунен усилвател
Артемизининът и неговите производни показват положителни имунни и противовъзпалителни ефекти ( R ). При лабораторни експерименти те биха могли да инхибират продуцирането на IL-1 (3, IL-6 и IL-8 от синовиални клетки на пациенти с RA ( R ).
Ацетоновият екстракт показва най-голям инхибиторен ефект върху азотния окис (NO), простагландин Е2 (PGE2) и провъзпалителния цитокин ( R ), индуциран от липополизахарид .
Артемизининът инхибира симпатиковото ремоделиране след инфаркт на миокарда, вероятно чрез противовъзпалителен ефект ( R ).
Той също така потиска TNF-a експресията и Т-хелпери 1 (Th) / Th17 реакции в модел на колиби на TNB (възможна потенциална терапия за болестта на Crohn) ( R ).
Той може да повиши селективно функцията на Т-клетките както при нормални мишки, така и при мишки със сингенна трансплантация на костен мозък ( R ).
Установено е, че производните на Артемизинин значително подобряват лупусния нефрит, редуцират и потискат производството на TNF-алфа от перитонеални макрофаги ( R ).
Тези производни са съобщени за лечение на експериментален алергичен енцефаломиелит (EAE) в добре установени миши модели чрез регулиране на равновесието между ефекторни Т клетки и регулаторни Т клетки ( R1 , R2 , R3 ).
Artesunate подобрява експерименталното възпаление на алергичните дихателни пътища. Могат да бъдат полезни при лечението на алергична астма ( R1 , R2 ).
Възможни нежелани реакции
Страничните ефекти са подобни на симптомите на малария: гадене, повръщане, анорексия и световъртеж ( R ).
Рядък, но сериозен нежелан ефект е алергичната реакция ( R ).
Лечението с Артемизинин може да доведе до разкъсване на червените кръвни клетки, особено при тези с предразполагащи фактори за анемия ( R ).
Други възможни нежелани странични ефекти могат да възникнат от комбинирана терапия (ACT), а именно хепатит и забавена хемолитична анемия ( R , R 2).
Михаил Христов е завършил Университетска химия, специалност Органик-химик, в ПУ „Паисий Хилендарски“ (1986-1991г.). След това е специализирал „Храни, хранене и диетика“ в УХТ Пловдив (2003-2005 г.).
Започнал като ревностен последовател на класиците на здравословния начин на живот (Пол Браг и др.), Михаил Христов хармонизира непрекъснато своите разбирания със съвременните научни открития и подходи – както в теоретичен, така и в практичен план. Той работи неуморно за популяризирането на тези важни за живота принципи, залагайки основно на ортомолекулярната (клетъчна) нутрициология.
Михаил Христов изнася редовно лекции на тема „Хранене при бременност и кърмене“ (в Центъра за бременни жени – гр. Пловдив), както и лекции и беседи по Здравословно хранене и начин на живот (по фестивали и други събития), участва в научни публикации, радио- и телевизионни предавания, разработване на диетични храни и менюта, и други проекти. Член е на Българското научно дружество по хранене и диететика и участва със свои колеги в доклади пред конгреси и конференции на Дружеството.
По повод на нашумяли напоследък публикации в пресата по темата за полезните лечебни свойства на сладкия пелин, известният пловдивски диетолог Михаил Христов реши да се обърне към един от специалистите в областта на природната медицина, храненето и диететиката, който препоръчва сладкия пелин като антивирусна профилактика и естествена подкрепа за имунната система – Д-р Владимир Шишков.
Д-р Владимир Шишков
Д-р Владимир Шишков завършва медицина във ВМИ – Пловдив през 1990 г. От самото си завършване работи активно в областта на фармацията, храненето, лечението и превантивната медицина както с природни, така и с фармацевтични продукти, храни и хранителни добавки. Член е на Българското научно дружество по хранене и диететика.
Д-р Шишков е профилиран в изграждането на цялостни индивидуални лечебно-профилактични и оздравителни програми, както и на диетични планове, насочени към преодоляване на здравословни проблеми и достигане на състоянието на “оптимално здраве”, в съответствие с принципите на орто-молекулярната медицина. Автор е на редица публикации в популярни издания, посветени на рационалното хранене и модерните хранителни добавки с доказан клиничен ефект и безопасното им приложение.
Михаил Христов : Защо растенията от рода Artemisia са толкова използвани в различните древни лечебни практики?
Д-р Шишков : Защото притежават изключителни лечебни свойства – за повече от 150 вида болести, свързани с различни едри паразити, бактерии, вируси, гъбички; храносмилателна система, черен дроб, жлъчка, панкреас, треска, настинки, малария, грип, сърцебиене, кръвно налягане, мускулна слабост, различни инфекции, кожни заболявания, страхова психоза, тревожност, проблеми с цикъла при жените, някои форми на безплодие, очни инфекции и т.н.
Михаил Христов : Какви видове пелин в диво състояние се срещат в България?
Д-р Шишков :В България се срещат диворастящи около 25 и повече вида пелин . Това са Обикновен (черен) див пелин (Artemisia vulgaris) ;Бял (абсентов, горчив) пелин (A. absinthium) ; Полски пелин (A. campestris) ; Метловиден пелин (A. scoparia) ; Бял пелин (катраника) (A. alba) ;Морски пелин (A. maritima) ;Австрийски пелин (A. austriaca) ;Черноморски (понтийски) пелин (A. pontica); Добруджански пелин (A. pedemontana); Скален пелин (A. eriantha), Дребнолистен пелин (A. chamaemelifolia)
Но според НАРЕДБА-47 за Хранителните добавки са забранени да се използват като субстанции, защото могат да бъдат опасни.
Михаил Христов : Защо именно вида Сладък пелин (Artemisia annua) е толкова подходящ за подобряване на здравето и профилактика при различни заболявания?
Д-р Шишков :Основното предимство на Сладкия пелин е ниската му токсичност и възможност за безопасно широко приложение дори и в доста високи концентрации. Сладкият пелин може широко да се използва като хранителна добавка. Освен това Сладкият пелин е единственият пелин, който може да бъде влаган в хранителни добавки, съобразно действащото законодателство.
Проучването показва, че Сладкият пелин има по-добър профил на безопасност от хлорохина и не по-малък превантивен ефект върху SARS-CoV-2.
Държа да кажа преди всичко, че препоръчвам приема на сладък пелин като превенция срещу вирусни инфекции, подчертавам това, включително и COVID-19. Да не забравяме, че хидроксихлорохина не е достъпен за всеки и се прилага само под лекарско наблюдение, докато Сладкият пелин може да се намери и не е скъп.
Разполагам и с друго интересно проучване, което показва, че активните вещества в Сладкия пелин са ефективни и срещу няколко други вируса, вкл. HIV и HCMV.
Сладкият пелин е най-добрият заместител на хлорохина.
Спомням си как още в самото начало на пандемията, китайските лекари съобщиха за първите успешни резултати при използването на хлорохин (който всъщност, е синтетичен аналог на хинина), а хининът е може би най-известното в цял свят антималарийно средство. Оказва се обаче, че пелинът също има доказано мощен антималариен ефект. И двете билки, по някакъв вероятно много сроден механизъм, блокират достъпа на маларийния плазмодий до хемоглобина в еритроцитите.
Оттук- нататък логиката е проста – ако успеем да блокираме достъпа на вируса до хемоглобина, ще предотвратим в много голяма степен пораженията, които той би нанесъл в микроциркулацията и свързаните с тях органни увреждания. И след като въпросното проучване демонстрира ефективността на пелина (и дори неговото превъзходство над хидроксихлорокина) в това отношение, смятам че той спокойно би могъл да се използва в много добре преценена, от гледна точка на безопасност и ефективност, доза за превенция на заболяването COVID-19, поне за известен период от време. Да не забравяме също, че сладкият пелин е достъпна и не особено скъпа хранителна добавка. Дори мои колеги конвенционални лекари повярваха, щом видяха сериозното проучване и самите те започнаха да приемат Сладък пелин в оптимална доза.
Михаил Христов : Какъв е произхода на този вид пелин?
Д-р Шишков : Сладкият пелин се смята, че произхожда от умерения пояс на Азия (Китай и др.).
Михаил Христов : Какви са научните данни за неговите многобройни ползи за здравето и при какви състояния се препоръчва?
Д-р Шишков :Научните данни са огромни в момента и не може да се отговори накратко. Бих искал да спомена, че в момента науката преоткрива колелото с пелините и заради китайските разработки (вкл. и една Нобелова награда) вниманието е насочено главно към сладкия пелин и неговото култивиране. Въпреки сериозните биохимични изследвания все още огромна група от активни вещества остават неизвестни и непроучени в сладкия пелин. Бизнес интересът разбираемо е фокусиран върху главната активна субстанция – артемизинин, но според древните фитотерапевтични школи и модерните фармакологични схващания, билките следва да се прилагат в своята цялост, тъй като сами по себе си са една биохимична „екосистема” и по този начин ще се минимизират вредните странични ефекти, реакцията на организма ще бъде адекватна и хармонична и лечебните ефекти трайни.
Михаил Христов : Коя е оптималната форма и дозировка за приложение на Сладкия пелин и при какви състояния го препоръчвате?
Д-р Шишков :Според моята практика най-оптималната форма за прилагане е смляна качествена сушена дрога (листа) на прах, която може да се използва в капсули. Дозировката, която прилагам е 350мг без никакви оплаквания и регистрирани до момента от мен странични ефекти.
Разбира се, за масово лесно, бързо и достъпно приложение най-удобни са стандартизираните изсушени билки в капсулни форми.
Как може да сме сигурни, че конкретен продукт от Сладък пелин е с най-високо качество и отговаря на най-високите стандарти за безопасност и гарантира най-много ползи за здравето?
Всичко зависи от нивото на качествен контрол който съответния производител прилага при отглеждането, обработката и високите качествени стандарти прилагани при капсулирането на изсушената билкова смес. Най-висококачествен е био-пелинът, който се капсулира във фармацевтични предприятия покриващи лекарствените стандарти.
Михаил Христов : Може ли сами вкъщи да си приготвяме сладкия пелин за тази цел? Как да го приемаме?
Д-р Шишков: В момента на пазара в България се предлагат няколко вида продукти, съдържащи разрешения за употреба в хранителни добавки вид пелин – Artemisia annua (сладък пелин), основно под формата на сушена билка в капсули. Аз лично не бих препоръчал да се употребява директно откъсната билка или билкова отвара от пелин, освен ако приемът не е предписан и контролиран от опитен фитотерапевт или лекар. Проблемът е, че активните вещества в сладкия пелин могат да варират в големи граници, в зависимост от начина на приготвянето на билката и многобройни други фактори на средата. Значително по-безопасно е да се приема стандартизираната билка в подходяща дозировъчна форма. Клиничните проучвания са правени върху билков екстракт, а не върху сушена билка (в която активните вещества са в доста по-ниска концентрация), показват, че оптималната дневна доза варира около 300-400 мг дневно, при липса на противопоказания.
Михаил Христов : Има ли вече на българския пазар продукти, които да отговарят на описаните в предния въпрос високи стандарти?
Д-р Шишков : Да, един такъв продукт например е Сладкият Био-пелин на компанията Ахрида Био .
Михаил Христов : Вие лично ползвате ли въпросния продукт, препоръчвате ли го на вашите пациенти и ако да – за какви цели?
Д-р Шишков :Да, в момента приемам по 1 капсула дневно именно от този продукт, като част от моят интегрален природен протокол за имунопрофилактика на вирусни инфекции. Всеки може да се запознае с протокола на моята професионална страничка във Фейсбук, както и да ми зададе своите въпроси по тази и други теми, свързани със здравословното хранене и здравословен начин на живот.
Studies Show Artemisia annua (Plant) Kills 98% of Cancer Cells in 16 Hours.
MARCH 10, 2014
Според изследвания, публикувани в списание „Life Science“, Aртемизининът, производно на „Sweet Wormwood“ или „Artemisia Annua“ се използва в китайската медицина и може да убие 98% от раковите клетки на белите дробове за по-малко от 16 часа.
Използваната билка самa по себе си намалява броя на раковите клетки на белите дробове до 28%, но в комбинация с желязо „Artemisia Annua“ успешно и напълно „изтрива“ рака, а в експеримента тази билка не оказва влияние върху здравите бели дробове.
Артемизининът в миналото е бил използван като мощен антималариен лекарствен продукт, но сега е доказано, че това лечение също е ефективно в борбата с рака.
Когато учените добавят желязо, докато провеждат изследването, което по-късно се отнася към белодробната тъкан, особено към клетките, заразени с рак, Aртемизининът селективно атакува „лошите“ клетки и оставя „добрите“ клетки недокоснати.
„По принцип нашите резултати показват, че Aртемизининът спира транскрипционния фактор“ E2F1 „и се намесва в унищожаването на раковите клетки на белия дроб, което означава, че той представя транскрипционния начин, според който Aртемизининът контролира растежа на репродуктивния раков клетъчен растеж“, се посочва в заключението на изследванията, проведени в ракова лаборатория в Калифорнийския университет.
Списанието „Life Science“ съобщава за проучване на университета във Вашингтон, проведено от д-р Наренда Сингх и Хенри Лай. Досега това е най-широкото проучване, проведено в САЩ за Aртемизинин.
Проучването е проведено в лаборатория в тъканни проби (in vitro), в съпоставка с проучване върху животни или хора (in vivo). Използвайки резистентно на радиация разнообразие от клетки на рак на гърдата (които също имат висока склонност към натрупване на желязо), Aртемизининът причинява 75% смърт на раковите клетки след само 8 часа; и почти 100 процента са унищожени само за 24 часа. Оригиналното изследване може да се намери тук.
Желязото се натрупва в раковите клетки чрез специални рецептори, които помагат при клетъчното делене. Нормалните клетки също имат тези рецептори, но раковите клетки ги имат в по-големи количества и в съответствие с това, раковите клетки могат да бъдат целева комбинация от желязо и Aртемизинин.
Има многобройни експерименти, проведени досега, и всички те доказват, че в комбинация с желязо Aртемизининът може ефективно да унищожи рака и това познание е било използвано в Китай от хиляди години за лечение на малария.
Маларийният паразит не може да оцелее в присъствието на Aртемизинин, защото е богат на желязо, а биоинженерите Хенри Лаи и Нарендра Синг от Университета във Вашингтон са първите учени, които откриха това. Оригиналната статия, публикувана в списанието, може да се намери тук , където учените заключават, че „Артемизининът и арттемизинин-маркирани желязосъдържащи съединения могат да се превърнат в мощни противоракови лекарства“.
Тук има още едно проучване, извършено от партньорски екип, което може да се прочете в линка.
Противоракови свойства на производни на Артемизинин и тяхната целева доставка чрез конюгиране с трансферин.
Артемизининът и неговите производни са добре известни антималарийни лекарства и особено полезни за лечението на инфекция на паразити на малария от Plasmodium falciparum, резистентни към традиционните антималарийни лекарства. Артемизининът има ендопероксиден мост, който се активира от интрапаразитно хеме-желязо, за да образува свободни радикали, които убиват маларийните паразити чрез алкилиране на биомолекули. През последните години има много съобщения за противоракови действия на Артемизинин както in vitro, така и in vivo. Artemisinins имат инхибиторни ефекти върху растежа на раковите клетки, включително и към много ракови клетъчни линии, устойчиви на лекарства и радиация. Цитотоксичният ефект на Артемизинина е специфичен за раковите клетки, тъй като повечето ракови клетки експресират висока концентрация на трансферинови рецептори върху клетъчната повърхност и имат по-висок приток на железни йони, отколкото нормалните клетки чрез трансферинов механизъм. В допълнение, някои аналози на Артемизинина са показали, че имат антиангиогенезна активност. Артемизининът, маркиран с трансферин чрез въглехидратна верига, също е доказано, че има висока активност и специфичност срещу раковите клетки. Конюгирането позволява целева доставка на Артемизинина в ракови клетки. В този преглед се обсъждат противораковите действия и механизмите на действие на Артемизинините и трансфериновия конюгат.
Изследователският професор Хенри Лай и помощник-професорът Нарендра Сингх са използвали химичните свойства на екстрактите от Artemisia за прицелване на клетки от рак на гърдата с изненадващо ефективни резултати. Проучване в изданието от 2001 г. на списание Life Sciences описва как екстрактът убива практически всички човешки ракови клетки от рак на гърдата, изложени на него в рамките на 16 часа.
Изследователският професор Хенри Лай и помощник-професорът Нарендра Сингх са използвали химическите свойства на екстрактите от Артемисиа за третиране на клетки от рак на гърдата с изненадващо ефективни резултати. Проучване в изданието от 2001г. на списание Life Sciences описва как екстрактът убива практически всички човешки ракови клетки при рак на гърдата, изложени на него в рамките на 16 часа.
„Не само изглежда, че е ефективна, но е много селективна. Това е силно токсично за раковите клетки, но има незначително
въздействие върху нормалните клетки на гърдата.“ Лай
Artemisia annua: старо лечение за малария – ново лечение за рак
Още през 60-те години на миналия век, по призив на председателя Мао, китайците отделят много време и енергия за разследване на лечебни растения, в желанието си да се справят без западната медицина. От този момент до сега химическият екстракт от растението, наречен Artemisinin, е изследван и използван терапевтично за лечение на малария. Изглежда, че е значително по-ефективен при убиването на паразита, причиняващ малария, отколкото някое от наличните фармацевтични лекарства. Световната здравна организация понастоящем насърчава производството и употребата на тези екстракти като ново, по-ефективно и по-евтино лечение. Извлеченията от Artemisia са се оказали евтини и безопасни.
Двама биоинженери изследователи във Вашингтонския университет са направили още една крачка напред в използването на Артемисия.
Изследователският професор Хенри Лай и помощник-професорът Нарендра Сингх са използвали химичните свойства на екстрактите от Артемисия за атакуване на клетки от рак на гърдата с изненадващо ефективни резултати. Проучване в изданието от 2001 г. на списание Life Sciences описва как екстрактът убива практически всички човешки ракови клетки от рак на гърдата, изложени на него в рамките на 16 часа.
„Не само изглежда, че е ефективна, но е много селективна“, каза Лай. „Това е силно токсично за раковите клетки, но има незначително въздействие върху нормалните клетки на гърдата.“
Съединението помага за контролиране на маларията, защото реагира с високите концентрации на желязо, открити в маларийния паразит. Когато Артемизининът влиза в контакт с желязо, настъпва химическа реакция, при която се отделят атоми, които химиците наричат “свободни радикали“. Свободните радикали атакуват клетъчните мембрани, разрушават ги и убиват едноклетъчния паразит.
Преди около седем години Лай започна прави хипотеза, че процесът може да работи и при рак.
„Раковите клетки се нуждаят от много желязо, за да възпроизвеждат своето ДНК, когато се делят“ – обяснява Лай. „В резултат на това раковите клетки имат много по-високи концентрации на желязо в сравнение с нормалните клетки. Когато започнахме да разбираме как работи Артемизининът, започнах да се чудя дали можем да използваме това знание, за да насочим към раковите клетки.“
Лай разработва потенциален метод и започна да търси финансиране, като получава субсидия от „Фонда за рака на гърдата“ в Сан Франциско. Междувременно Университетът във Вашингтон (UW) патентова идеята.
В основата на идеята, според Лай и Сингх, е да се „напомпят“ раковите клетки с максимални концентрации на желязо, след това да се въведе Артемизинин, който избирателно убива рака.
Само след 8 часа, 25% от раковите клетки оцеляват. След 16 часа, почти всички клетки били мъртви.
По-ранно проучване, включващо левкемични клетки, дава още по-впечатляващи резултати. Тези клетки се елиминират в рамките на 8 часа. Възможно обяснение може да бъде нивото на желязо в клетките на левкемията.
„Те имат една от най-високите концентрации на желязо в раковите клетки“, обясни Лай. „Левкемичните клетки могат да имат повече от 1000 пъти повече концентрацията на желязо от нормалните клетки.“
Следващата стъпка, според Лай, е тестването на животни. В тази област са направени ограничени тестове. В по-ранно проучване върху куче с рак на костите, толкова тежко, че не може да ходи, се възстановява напълно 5 дни след получаване на лечението.
При повече от 800 статии в базата данни на PubMed за Artemisia, само около 30 се занимават с рак. От тях никой не описва клиничните изпитвания на Artemisia при хората. Тогава започва да се разглежда като терапия с голям потенциал. Няколко лекари съобщават за анекдотична полза при лечението на пациенти с рак, но внимателното изследване на повечето от тези истории ги намира за неубедителни. Има и други терапии прилагани съвместно, които могат да доведат до предполагаемите ползи. Това е една от тези терапии, които показват голямо обещание, но в този момент не е ясно дали работи с рак. Изследванията продължават и повече тестове участват в анекдотичната база данни за Artemisia.
Защо растенията от рода Artemisia са толкова използвани в различните древни лечебни практики?
Защото притежават изключителни лечебни свойства – за повече от 150 вида болести, свързани с различни едри паразити, бактерии, вируси, гъбички; храносмилателна система, черен дроб, жлъчка, панкреас, треска, настинки, малария, грип, сърцебиене, кръвно налягане, мускулна слабост, различни инфекции, кожни заболявания, страхова психоза, тревожност, проблеми с цикъла при жените, някои форми на безплодие, очни инфекции и т.н.
Лечебно приложение е широко спектърно, прилага се срещу:
при подагра; при вялост, умора, изтощение (въбужда нервната система);
при злоядство, за възбуждане на апетита,
при диспепсия; ефективен срещу анорексия и булимия ;
за детоксикация и като антидот за различни природни и химически отрови;
за прочистване на кръвта, лимфата, жлъчката и храносмилателната система
за регулиране менструалния цикъл и за различни женски болести – аменория, менорагия, кисти, миоми, ендометриоза
регулира/модулира кръвното налягане , както и забавя и регулира пулса (при някои видове тахикардия, брахикардия);
различни болести на храносмилателната система – лениви черва, забавена перисталтика, лошо храносмилане, болест на Крон и проблеми/дисфункции на чревния микробиом,
при кандидози; много ефективен при кървави диарии и различни кръвоизливи и кръвотечения;
при всякакви форми на мускулна слабост (възстановяване на мускулния тонус), безпокойство, тревожност, депресия;
антисептично и антиспазмолитично действие;
някои форми на безплодие при жените и мъжете (вероятно свързани с паразити и инфекции); при всякакви възпаления на половите органи и някои венерически болести (гонорея, частично при сифилис); комплексно антибиотично действие (вирусни, бактериални, гъбични инфекции);
срещу почти всякакви инфекции и гнойни процеси;
ставни заболявания, ревматизъм, различни видове артрити;
разнообразни чернодробни болести (хепатит В, цироза и др., като хепатопротектор);
за изхвърляне на камъни в жлъчката (особено, когато са холрестеролови);
прочиства от отлагания кръвоносната система (включително и венозната) и лимфата; укрепва стените на капилярите и вените; защитава, възстановява и укрепва миелиновите обвивки на нервите
Кои са растенията от рода Artemisia ?
В България са познати като пелин. Тук се срещат над 25 диворастящи вида – Обикновен (черен) див пелин (Artemisia vulgaris) ;Бял (абсентов, горчив) пелин (A. absinthium) ; Полски пелин (A. campestris) ; Метловиден пелин (A. scoparia) ; Бял пелин (катраника) (A. alba) ;Морски пелин (A. maritima) ;Австрийски пелин (A. austriaca) ;Черноморски (понтийски) пелин (A. pontica); Добруджански пелин (A. pedemontana); Скален пелин (A. eriantha), Дребнолистен пелин (A. chamaemelifolia)
Но според НАРЕДБА-47 за хранителните добавки са забранени да се използват като субстанции, защото могат да бъдат опасни, тъй като токсичността в тях е висока.
Кой вид пелин се използва за лечението на изброените горе заболявания ?
Единственият разрешен за използване пелин според Наредба-47 на законодателството това е Artemisia annua – така нареченият Сладък пелин. За жалост , той не се среща свободно в България , родината му е Китай. От години Сладкият пелин предизвиква интересът на лекари и учени на по цял свят и е обект на множество изследвания. Проучваният му дори донесоха Нобелова награда за физиология и медицина от 2015 г. на д-р. Юю Tу, Уилям С. Кембъл и Сатоши Омура.
Защо именно вида сладък пелин (Artemisia annua) е толкова подходящ за подобряване на здравето и профилактика при различни заболявания?
Основното предимство на сладкия пелин е ниската му токсичност и възможност за безопасно широко приложение дори и в доста високи концентрации. Сладкият пелин може широко да се използва като хранителна добавка от всякакъв вид. Освен това сладкия пелин е единственият пелин, който може да бъде влаган в хранителни добавки, съобразно действащото законодателство.
Коя е оптималната форма и дозировка за приложение на Сладкия пелин/Артемизинин/ ?
Най-оптималната форма за прилагане е смляна качествена сушената билка (листа) на прах, която може да се използва в капсули. Дозировката, е 350мг без никакви оплаквания и регистрирани странични ефекти. Сладкият пелин не се приемам във вид на час, защото при нагряване, се изпарява Артемизинина от растението и полезните свойства изчезват.
Как може да сме сигурни, че конкретен продукт от Сладък пелин/Артемизинин/ е с най-високо качество и отговаря на най-високите стандарти за безопасност и гарантира най-много ползи за здравето?
Най-важното е да сме сигурни за произхода! За съжаление на пазара е пълно с множество вносни билкови препарати, за които не можем да сме сигурни дори какво съдържат, а какво да говорим за качеството на субстанциите. Най-висококачествен е био-пелинът който се капсулира във фармацевтични предприятия покриващи лекарствените стандарти и може да се проследи целият процес на отглеждане , до краен продукт.
Има ли вече на българския пазар продукти, които да отговарят на описаните в предишния въпрос високи стандарти?
Един такъв продукт например е сладкия био-пелин на компанията ahrida Bio . Повече от 5 години те се стремят да предлагат Сладък пелин от най-високо качество, който успешно се конкурира с големи компании и в чужбина. Те са култивирали билката и я отглеждат в екологично чист район на България ( Източните Родопи). Използват само листата на билката, в продукта си , които са силно наситени на Aртемизинин. Притежават и сертификат за био производство. Билките са полезни и ефикасни, само ако се отглеждат правилно, затова е важно да знаем произхода на субстанциите, а не само фирмата, която си капсулира.
В какъв вид предлагат Сладкият пелин/Артемизинин/?
Изследванията сочат, че най-оптимален ефект се постига при прием на 350мг на ден в продължение на 3 месеца , от качествена изсушена билка. Капсулите Артемизинин от ahrida BIO са с точната дозировка от 350мг и опаковката съдържа 90 капсули , които да стигат за пълния курс от 3 месеца.
Къде могат да се открият капсулите Артемизинин от ahrida BIO ?
Те имат сключен договор за дистрибуция с една от големите вериги аптеки – ФРАМАР. От там може да бъде закупен продукта или директно от тях , през онлайн магазина им – www.ahridabio.com или на телефон 0886153636 . Доставят го до 1-3 работни дни с безплатна доставка .
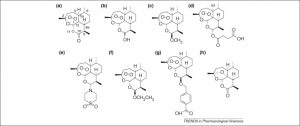
Забележителната история за откриването на Aртемизинина ( Фигура 1а ) и установяването на неговата антималарийна активност от китайски учени представлява едно от големите открития в медицината през втората половина на 20 век [1] . Чрез колективно усилие, наричано общо „Проект 523“, бе произведен китайският дихидроартемизинин (DHA; Фигура 1b ), артеметер ( Фигура 1 с) и артестенат ( Фигура 1d ) през 70-те години. Това са тези производни [с други, включително артемизона ( фигура 1 е), артеинер ( фигура 1 е) и артелинова киселина ( фигура 1ж), общоизвестни като „артемизинини“], които сега имат решаващ принос за борбата с маларията, една от най-унищожителните инфекции. Мащабът на проблема с маларията годишно засяга 500 милиона случая. Този очарователен клас лекарства, със структури, които са толкова различни от класическите хинолинови антималариали, са особено ценни, когато се използват в комбинация с други антималарии [2,3] .
Фигура 1
Химични структури на артемизинините. Artemisinin (a), изолиран в кристална форма през 1973 г. от Artemisia annua и производни на дихидроартемизинин (DHA) (b) , artemether (c) , artesunate (d) и arteether (f), бяха създаден за първи път от китайски учени през 70-те [1] . Artemisone (e) , представител на новия клас артемизинин, известен като амино-артемизинини, е лечебен при клинични проучвания при една трета от дозовия режим на артесунат. Характеризира се с ниска токсичност [56] . Artelinate (g) е създаден в Института за изследване на екипа на Уолтър Рийд ( http://wrair-www.army.mil), но е оттеглено поради опасения за токсичност [112] . Деоксиартемизинин (h) , който няма пероксиден мост, е биологично инертен.
Артемизинините също са били подложени на проучвания, насочени към проучване на други употреби на този клас лекарства. Артемизинините са активни срещу други паразитни видове in vitro , включително протозои, които са филогенетично несвързани с паразити на апикомплексан, такива като вида Plasmodium, които причиняват малария. Артезизините също действат срещу метазоански паразити като Schistosoma spp. Техните анти-болестни свойства включват мощна противоракова активност в in vitro проучвания и in vivoмодел на колоректален рак. Взети заедно с докладите за случаи, описващи ползите при различни ракови заболявания, наскоро е публикувано клинично проучване за краткосрочна употреба при рак на белите дробове, установените данни за безопасност при деца и възрастни с малария и ниските разходи, има убедителни причини да се проучи техният принос за лечение на тумори, които изискват терапии с адювантни и нео-адювантни средства. Този селективен преглед се фокусира върху бързо развиващите се области на науката и употребата на Артемизинин и илюстрира защо Артемизинините имат потенциала да се съревновават с ацетилсалициловата киселина в ширината на техните анти-болестни свойства.
Има сериозен дебат относно механизмите на антималариалното действие на Артемизинините. Ендопероксидният мост ( фигура 1 ) е в основата на антипаразитната активност на артемизинините, въпреки че химичният характер на взаимодействието между Артемизинините (особено ендопероксид) и паразитните мишени не е добре разбран. Ролята на желязото в антималариалните действия на Артемизинините също се обсъжда [4], защото тези катиони могат да катализират in vitro реакциите на някои artemisinins, включително тяхното разпадане във водни разтвори.
Един въпрос поставя акцент върху по-нататъшните дискусии: има ли една важна цел за artemisinins в Plasmodium spp. или има ли няколко цели? Напълно синтетичните триоксолани, които съдържат ендопероксиден мост, но нямат други характеристики на артемизинините, са увеличили сложността на дебата относно механизмите на действие на Артемизинините [5] . Много групи, включително и нашите, са прегледали последните събития [6-9], Изясняването на механизмите на действие на Артемизинините е важно за разбирането как структурно свързани лекарства, като например напълно синтетичните триоксолани, могат да работят и основа за развитие на резистентност от паразити към този клас антималарийни средства. Ясно е, че структурното преценяване на предполагаемите цели трябва да допринесе за проектирането на производни, които не са осакатени от мутациите в целевите групи, както се вижда от подходите, използвани при разработването на нови инхибитори на дихидрофолатна редуктаза [10,11] .
Малариите причинени от гризачи също са полезни модели за разбиране на възможните механизми на резистентност към различни класове антималарии [12,13] . Генетичните анализи, разрешени от инфекцията с Plasmodium chabaudi при мишки, идентифицират локус, свързан с резистентност към артемизинин, който е стабилен след преминаването на комари [14,15], Връзките с резистентността към артемизинин са стеснени до дебитабилизиращия ензим (между другото), който може да функционира в ендоплазмения ретикулум на паразитите и да участва в стресовия отговор. Други групи са установили стабилни щамове резистентни към артемизинин, което потвърждава, че резистентността към Артемизинин може да се развие чрез стандартни процедури за подбор, а не (за съжаление) е изключително рядко събитие и може да възникне и при повече от един механизъм [16-18] .
Молекулярни цели на артемизинините
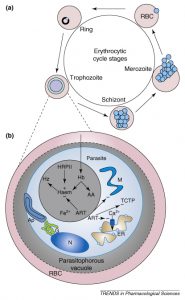
Plasmodium falciparum се умножава в червените кръвни клетки, а храносмилането на хемоглобина по време на 48-часовия животоспасяващ цикъл е от съществено значение за оцеляването на паразитите ( каре 1 ). В продължение на много години са предложени артемисинини да действат върху процесите на храносмилане на паразитен хемоглобин в рамките на „хранителната вакуола“ ( каре 1 , фигура I b). Други проучвания показват, че артезизините могат също така да бъдат насочени към паразитен митохондрион или транслационно контролирания туморен протеин (TCTP) и PfATP6, паразитно-кодирана саркоплазмично-ендоплазмена ретикулумна калциева ATPase (SERCA). Тези хипотези са разгледани по-подробно тук.
Диаграмата показва сложния жизнен цикъл на Plasmodium falciparum. Съкращения: АА, аминокиселини; Ap, апикопласт; ART, артемизинини; DV, храносмилателна вакуола; ER, ендоплазматичен ретикулум; G, Golgi апаратура; Hb, хемоглобин; Hz, хемозоин; М, митохондрион; N, ядро; RBC, червени кръвни клетки; TCTP, транслационно контролиран туморен протеин.
Клетка 1
Вътрееротроцитния паразит и предложените цели на артемизинините
Противопаразитните паразити, причиняващи човешка малария, имат сложни жизнени цикли, изискващи както вектори на комари, така и човешки гостоприемници с три цикъла на асексуални и един цикъл на сексуално възпроизвеждане. Една от асексуалните фази се осъществява в червените кръвни клетки на неговия гостоприемник ( Фигура Iа ). Инвазивните форми, наречени мерозоити, влизат в червените кръвни клетки и остават сравнително метаболитно неактивни (в сравнение с по-късните асексуални етапи на развитие) в продължение на 10-15 часа (етапа на пръстена). След това паразитът претърпява бърза фаза на растеж в следващите 25 часа (формиращ тропозоитния стадий), през което време паразитът усвоява по-голямата част от хемоглобина на клетката-гостоприемник и расте за да запълни> 50% от обема на клетката гостоприемник Хемоглобинът се усвоява в рамките на хранителна вакуола ( фигура 1)б), което води до образуването на хем. Тъй като се образува хем, той се свързва чрез една от периферните карбоксилни групи с Fe3 + на съседен хем, за да образува неразтворим хемозоин. Предложено е, въпреки че не е доказано, че този процес се подпомага от протеин, наречен протеин II, богат на хистидин. В края на фазата на тропозоида паразитът се дели няколко пъти (стадий на сризона) преди да се разхлади (около 48 часа след инвазията) клетката гостоприемник, за да се освободят новообразуваните мерозоти, които продължават цикъла.
Artemisinins, които може да не изискват активиране с Fe2 + , са предложени в продължение на няколко години, за да се насочат към няколко различни пътища ( фигура I b), включително пътя на детоксикация на гърба, митохондрион, TCTP и Ca 2+ помпа, локализирани към ендоплазмения ретикулум (наречен PfATP6).
Хаем пътека
Хемозоинът е паразитен пигмент, депониран в хранолна кутия ( кутия 1 ) след храносмилането на хемоглобина. Дълго се предлага като цел на артемизинините, въпреки че плазмодиалните етапи, които са най-чувствителни към активността на артемизинините, са твърде малки, за да проявят видим пигмент (прегледани в Реф. [19,20] ). Ендопероксидният мост на артемизинин се предлага да бъде активиран от желязо желязо, за да се генерират свободни радикали (на окси или С-центриран сорт) при in vitro експерименти и впоследствие да се алкилира хеми. Тъй като желязото е основният елемент, депониран в хемозоина, се препоръчва храносмилането на хемоглобин от паразити, за да ги направи податливи на убиване от локално активирани артемизинини.
Няколко проучвания за локализация обаче показват, че повечето артемисинин, взети в паразити, е извън техните вакуоли за храна [21,22] . Някои изследвания с флуоресцентни производни на артемизинин показват наличие на вакуоларна локализация на храната [23] , което може би представлява трафик на самия флуорофор. Този трафик на напълно синтетичен флуоресцентен антималариен триоксолан може също да обясни диференциални резултати от локализирането (един паразит със сигнал в цитозола, а другият в хранопровода за храна), наблюдаван за два паразита, които имат същия еритроцит [24] . Синтетичните триоксалани като ОЗ277 са по-крехки от полусинтетичните производни на артемизинин, когато се анализират във водни разтвори [4,25,26], и те също изглежда да се разграждат лесно в паразитирани еритроцити [27] . Тези свойства могат да повлияят на оценките на потентността.
Други доказателства за несъответствието на паразитния пигмент при действието на артемизинин идват от тяхната мощна активност срещу паразити, които не предизвикват пигмент апикомплексан (виж по-нататък). Съществуват също различия между някои in vitro анализи на хемо алкилирането с триоксолани и естествени и полусинтетични артезинини [25] . Наблюдаваната корелация между антималариалните ефекти на триоксоланите и тяхната склонност към алкилиране на хем [25] не се наблюдава при артемизинини, което означава, че тези класове антималарийни могат да имат различни начини на действие или че всъщност пътят на хем може да е без значение. Триоксолан ОЗ277 инхибира активността на PfATP6 калциевата АТФ-аза, когато се експресира в ооцити [24]при ниски (цМ) концентрации. Това може да се дължи на разлагане на съединението при условията на анализа или други аспекти на in vitro аналитичната система. Изследването на по-стабилните триоксолани може да разреши някои от тези проблеми. Съществува и корелация ( r 2 = 0.5, n = 38, p = 0.002) между паразитицидните активности на артесунат и ОЗ277, тествани срещу полеви изолати, без корелация между OZ277 и други класове антималарийни такива като хинолини [28], Тази корелация може да представлява обща (несвързана с целта) склонност на паразитите да бъдат податливи на ендопероксиди, но също така е в съответствие с хипотезата на споделената цел за механизми на действие, като PfATP6 е пример за такава цел.
Като вариант на хематологичната хипотеза, реакцията с богатия на хистидин протеин от паразити (HRPII; Box 1 ) може също да участва в антималарийната активност [29], тъй като HRPII спомага за усвояването на хемоглобин. Въпреки това, много малко HRPII се секретира в ранните стадии на кръвообращението ( Каре 1 ), които са най-податливи на артезинини [30,31] .
Разбирането на взаимодействията между хемоглобините и артезинините е усложнено от промени в статуса на желязо, свързани с хемоглобинопатиите. По-високите концентрации на свободно желязо в хемоглобин-Е-съдържащите и таласемичните еритроцити намаляват паразитицидните ефекти на артемизинините, когато се изследват in vitro [32] . Въпреки това, in vivo кинетичните изследвания, използващи биотестове на артестенат и неговия активен метаболит, DHA, показват приблизително десетократно по-високи плазмени концентрации при а-таласемични субекти, когато участъците под кривите време-концентрация са били оценени [33] и хемоглобин Е черта може да увеличи паразита клирънс от артезинини [34] . Въпреки тези разлики между in vitroактивността на артемизинини, свързани с хемоглобиновия статус на еритроцитите на гостоприемника, таласемията не е влиятелен ко-вариант в популационния фармакокинетичен анализ на ректалния артесунат, използван за лечение на инфекции с Plasmodium vivax или P. falciparum . В допълнение, антималариалните активности на артемизинин срещу паразити на P. falciparum, култивирани в присъствието на карбоксихемоглобин, са значително по-високи, отколкото в присъствието на окси-хемоглобин. Това увеличение на активността на артемизинин е неочаквано, ако Fe 2+ е важен при активирането на артемизинини, защото карбокси-хемоглобинът инхибира хемо-Fe2 +че хемоглобиновото желязо не играе роля при активирането на артемизинин за антималариална активност и че конкурентното разграждане на артемизинин с хемоглобин действително отслабва неговата антималарийна активност [35,36] .
PfATP6
Подкрепящите аргументи за PfATP6, ортологът P. falciparum SERCA, като цел за артемисинини, са били разгледани наскоро [9] . Доказателствата от трансфекция в паразити на ДНК, кодираща PfATP6, които имат променена чувствителност към някои artemisinins, ще осигурят подходящи генетични тестове за PfATP6 хипотезата (проучвания в процес), които са получили подкрепа от данни от полеви изолати. Едно интересно проучване от Френска Гвиана показва ясна връзка между мутации в PfATP6 и намалена чувствителност към артеметер, особено при позиция 769 (заместване на Ser769Asn) [37] . Паразити с Ser769Asn имали медиана IC 50стойност> 20 пъти по-висока за артеметер (показваща артеметерна резистентност) в сравнение с паразитите без тази мутация [9] .
Подробна методология за in vitro анализите, използвани в по-ранната публикация [37], е представена в последващ доклад [38] . Липсата на лабораторно адаптирана линия, носеща мутацията на Ser769Asn, е критикувана, въпреки че има добре познати „разходи за фитнес“ (т.е. способността на устойчивите паразити да се запазят при отсъствието на натиск върху лекарството) на някои мутации за резистентност към култивирани паразити , както е показано за мутации в P. falciparum мултирезистентен ген 1 ( pfmdr1 ) [38-40] . Лабораторно получените трансфектанти, носещи мутацията Ser769Asn, ще изяснят своята роля в резистентността към артемизинин, особено когато се комбинират с ex vivoанализи на податливостта към артемизинини с модела на Xenopusoocyte. Африканският изолат, носещ мутацията Ser769Asn, все още е чувствителен към DHA и данни за чувствителност към артеметер не са докладвани ( Таблица 1 ). Тези наблюдения показват, че различните производни на артемизинин пораждат различни инхибиторни профили, когато срещат PfATP6 със специфичен полиморфизъм от едно място [41] , както се обсъжда другаде [42] . Структурното моделиране на мутацията Ser769Asn се оказа трудно, тъй като регионът, съдържащ тази мутация, има относително ниска прилика с SERCA на бозайници (в сравнение с други функционално запазени области), чиято кристална структура е налице [43], Този регион не е свързан с мястото на свързване на тапсигаргин на SERCA на бозайници, което в PfATP6 също е хипотетично за приемане на артемизинини въз основа на мутационни изследвания след експресиране в ооцити [44] .
маса 1
Полиморфизъм в PfATPase6 гена и in vitro чувствителност към артезизини на Plasmodium falciparum
Мутацията другаде в полевите изолати (позиция 243 в PfATP6) намалява чувствителността към DHA, въпреки че данните са достъпни само от два изолати [41] . Мониторингът на полиморфизмите в PfATP6 (и всъщност други последователности на транспортера) и свързването на резултатите с фенотипове чрез оценяване на чувствителността към артемисинини е вероятно да бъде от голямо значение за целта за откриване на ранни признаци на резистентност към артемизинин ( Таблица 1 ). Например, увеличеният брой копия на гена за резистентност към много лекарства pfmdr1 модулира чувствителността на паразитите към artemisinins in vitro , въпреки че клиничната значимост на това наблюдение не е установена [45] .
Други цели
Последните проучвания с хлебна мая показват, че митохондриална мембрана потенциал може да бъде прекъснато от артемизинин, когато се отглежда в nonfermentable условия (т.е. . Когато въглеродни източници, като например глицерол или етанол не се метаболизират чрез гликолиза) [46]. Въпреки това, уместността на тези наблюдения към антималарийната активност на артемизинините е неясна, защото други експерименти показват, че са необходими по-високи концентрации (тМ) артемизинини, за да предизвикат резистентни отговори към артемизинини в дрожди [47] . Освен това, новият клинично тестван артемизонов дериват artemisone няма ефект върху митохондриалния мембранен потенциал, нивата на реактивните кислородни видове или инхибирането на респираторната верига в невронните клетъчни линии [48] .
TCTP ортологът на P. falciparum е идентифициран преди няколко години като протеин алкилиран чрез радиомаркиран артемизинин. Няма нови доказателства, които да подкрепят идеята за TCTP като цел за artemisinins. Полевите изолати, които имат променлива чувствителност към артеметер, не са свързани с полиморфизмите на последователностите в TCTP [37] . Нито проучванията с животински модели на устойчиви на артезинин паразити не подкрепят включването на ТСТП като целева група [15] .Отидете на:
Свойства на артемизинините
Антималарийна активност на артемизинините – клинични приложения
Using artesunate to treat severe malaria in adults has been emphasized in recent publications [49]. Parenteral artesunate (including intramuscular artesunate [50]) is easier to administer and is associated with fewer adverse effects (e.g. hypoglycaemia) when compared with quinine [51], the only other drug used in severe malaria. Mortality in adults is also lower with artesunate than with quinine. Intrarectal treatment with artesunate of children or adults who cannot take medicines by mouth and suffer from symptoms of malaria away from healthcare facilities has also been studied in large scale (Phase IV) studies that will be reported soon. Both safety and efficacy have been established in smaller studies [52,53], Въпреки това, дете, лекувано с много високи ректални дози артесунат (88 mg kg -1 общо в сравнение с препоръчваните 10-20 mg kg -1 ), наскоро почина поради вероятна токсичност [54] .
Любопитно е, че пероралният артеметер и DHA се използват по-често в формулировки с фиксирана доза, отколкото в артестена. Artesunate може да има по-благоприятни свойства както по отношение на стабилността, така и по отношение на лекотата на съвместна формулировка в сравнение с DHA и по отношение на нежеланите ефекти при животинските модели в сравнение с артеметер [55] . По-новите полусинтетични производни на артемизинин като артемизона ( Фигура 1 д) запазват безопасността, но повишават ефикасността и трябва да се изучават за изпълнение срещу модели на резистентност към артемизинин [56] .
Активност срещу Toxoplasma gondii и други патогенни апикомплексан паразити
Изучаването на чувствителността на неплазмодиалните апикомплексани към артемизинините предоставя нови терапевтични възможности и осигурява нови механични прозрения. Ако организмите в короната на еукариотната група са податливи на артемисинини, тогава най-простото механистично интерпретиране е, че те функционират по подобен начин срещу тези филогенетично свързани организми. Например, Toxoplasma gondii е малко по-трагиращ паразит, отколкото Plasmodium spp., Особено за изследвания, използващи генетични манипулации или технологии за изображения. Ранната работа показа, че токсоплазмата е податлива на артезинини, въпреки че изисква концентрации в микромоларния диапазон да убиват паразити ( онлайн допълнителна таблица S1 ). Сега проучванията показват, че T. gondiiмогат да бъдат убити с наномоларни концентрации на артемизон в in vitro модели и че TgSERCA (ортологът PfATP6) е податлив на инхибиране от артемизинин, когато се експресира в дрожди [57] . Освен това, артезинините предизвикват нарушения на калциевия метаболизъм при паразити, които имат функционални последици върху машините за инвазия, и те могат да се различават, ако паразитите се култивират в клетките гостоприемници или като свободни живи организми [58] . Тези открития независимо подкрепят хипотезата, че паразитните SERCAs са цели за артезинини (както in vivo, така и след хетероложна експресия). Те също така показват, че остатъкът от глутаминова киселина, предсказан в трансмембранен сегмент 3 на TgSERCA, е допустим за податливостта към артемизинин [44], в съответствие с предложението, направено тук, че други ключови остатъци в TgSERCA могат да модулират чувствителността към артемизинин.
Видовете Babesia са инферитроцитни паразити, носещи кърлежи, които могат да инфектират хора в допълнение към различни домашни животни, в зависимост от вида на паразита. За разлика от плазмоидните инфекции, бабесията не генерира паразитофорен вакуол и не усвоява хемоглобин за хемозоин [59] . Въпреки това, някои видове също са податливи на убиване от артемизинин ( онлайн допълнителна таблица S1 ), като отново правят пътя на хемаглобин по-малко убедително за техните механизми на действие. Други свързани паразити имат променлива чувствителност към артезинини ( онлайн допълнителна таблица S1). Тези изследвания също установяват, че нито хемозоинът, нито хемоглобинът са от решаващо значение за антипаразитната активност на артемизинините. Ще бъде от интерес да се тества хипотезата за SERCA за механизма на действие на артемизинините в тези свързани патогенни паразити.
Дейност срещу други протозойни и метазоанови паразити
Artemisinins също са активни срещу филогенетично несвързани паразити, като едноклетъчните кинетопластиди и метазоанските хелминти ( онлайн допълнителна Таблица S2 , ефикасността срещу Schistosoma spp. Е разгледана другаде [60] ). Както алуминиеви (африкански), така и стеркорационни (американски) трипанозоми могат да бъдат убити с микромоларни концентрации на артемизинини, което показва, че артезинини могат да бъдат използвани като води, върху които да се оптимизират по-мощни производни [61] . Leishmania spp. също се убиват с микромоларни концентрации на артезинини ( онлайн допълнителна таблица S2 ). Тъй като тези инфекции обикновено се пренебрегват в портфейлите за разработване на лекарства, би било жалко, ако са обещаващиin vitro дейностите не се изследват по-задълбочено в съответните in vivo модели, вероятно използвани в комбинация с текущите терапии.
За метазоновите инфекции, по-специално за Schistosoma spp., Артеметер и артесунат са показали полезни дейности в изследвания на хора и модели на инфекция [60,62] . Първоначално идентифицирани в китайски проучвания [63] , тези наблюдения са обхванати и от африканските инфекции. Ограниченото портфолио от активни трематоцидни съединения засилва потенциала на артезизинин при лечението на Schistosoma mansoni и Schistosoma haematobium .
Антитуморни свойства на артемизинините
От края на 80-те години, противораковите свойства на артемизинините са изследвани in vitro ( онлайн допълнителна таблица S3 ). След по-подробни проучвания се установи, че артезинини като артесунат са активни срещу редица несвързани туморни клетъчни линии, от най-често срещаните видове като рак на дебелото черво, гърдата и белия дроб до левкемии и рак на панкреаса [64,65] . Проучванията също са идентифицирали потенциални общи механизми, като например нормализиране на възходящия път на Wnt / β-катенин при колоректален карцином [66] . Други пътища за противоракова активност включват инхибиране на засилена ангиогенеза свързана с тумори [67-77], Artemisinins инхибират пролиферацията, миграцията и образуването на епруветки на човешки ендотелни клетки от пъпна вена (HUVEC), инхибират свързването на повърхностните рецептори с HUVEC на съдовия ендотелен растежен фактор (VEGF) и намаляват експресията на VEGF рецепторите Flt-1 и KDR / flk-1 върху HUVECs [ 74, 75, 77] . В раковите клетки artemisinins намаляват експресията на VEGF рецептора KDR / flk-1 в туморните и ендотелните клетки и бавния растеж на HO-8910 ксеноприсадки на човешки рак на яйчника в голи мишки [67-69,75,77] . HUVEC апоптозата от artesunate се свързва с downregulation на Bcl-2 (B-клетъчна левкемия / лимфом 2) и с повишаване на регулацията на BAX (Bcl-2-свързан X протеин) [78] .
mRNA експресията на 30 от 90 свързани с ангиогенезата гени корелира значително с клетъчния отговор на артемизинини [70] . В този микрочип панел, има много основни ангиогенни регулатори, кодирани от гени като VEGFC , фибробластен растежен фактор-2 ( FGF2 ), матрична металопротеиназа-9 ( ММР9 ), тромбоспондин-1 ( THBS1 ) и хипоксия-индуциращ фактор α ( HIF1A). Фактът, че чувствителността и устойчивостта на туморните клетки може да се предвиди от иРНК нивата на експресия на ангиогенеза, свързани с гени, показва, че artemisinins разкриват техните противотуморни ефекти, поне отчасти, чрез инхибиране на туморна ангиогенеза. Свръхекспресия на ензими, свързани с модулация на оксидативния стрес като glutamylcysteine синтетаза, глутатион S-трансферази и ендотелен рецептор на растежния фактор за намаляване на чувствителността на туморните клетки към artemisinins [79,80], Важно е, че свръхекспресия на гените, кодиращи транспортери, които медиират лекарствена резистентност (например множествена лекарствена резистентност ген 1, множествена лекарствена резистентност свързан протеин 1 и рак на гърдата се протеин за резистентност), дихидрофолат редуктаза и рибонуклеотид редуктазата, които също предоставят резистентност към определени антитуморни лекарства, не засягат чувствителност, което показва, че артемизинините функционират по различен начин спрямо класическите химиотерапевтични средства за рак. Тези in vitro изследвания също така показват, че при някои ракови линии, доставката на желязо, например чрез използването на холотрансферин, повишава противораковите свойства на артемизинините [65,81-87] .
Ако артемизинините останат извлечени в голяма категория съединения, които имат интересни in vitro свойства срещу ракови заболявания, но не са изследвани достатъчно, за да оправдаят по-обширни клинични проучвания? Вероятно не, поради много причини. Първо, artesunate е евтино, безопасно, лесно администрирано и орално бионалично съединение, което действа при цели, различни от тези на много настоящи химиотерапевтични средства за рак, и е малко вероятно да взаимодейства неблагоприятно със съществуващите противоракови интервенции (P. Folb, лична комуникация). Второ, изследването на животински модел, носещ клетъчна линия на човешки колоректален рак, потвърждава, че артестенатът има независима антитуморна активност и може да свие първичните тумори и да намали риска от развитие на чернодробни метастази [66], В допълнение, проведените при хора проучвания на отделни случаи [88,89] , в допълнение към наскоро публикувано проучване Фаза ІІ на рак на белия дроб [90] , подкрепят бързото прилагане на проучвания на артесунат като първична или допълнителна антитуморна интервенция, особено за рак на дебелото черво и за левкемия (както се подкрепя от резултатите в онлайн допълнителната таблица S3 ).
Други потенциално полезни свойства на артемизининови съединения
В in vitro проучвания, няколко групи съобщават, че артемизинините имат антивирусни свойства. Artemisinins намаляват скоростите на репликация на вирусите на хепатит В и С [91,92] , редица човешки херпесни вируси [93-95] , HIV-1 [96] , грипен вирус А [93,97] и говежди вирус на диария [ 98] в ниския микромоларен диапазон. Artesunate също е ефективен при намаляване на броя на копитите на CMV (човешки херпес вирус 5) при имуносупресирано 12-годишно дете [99] и е използван (100 mg дневно, перорално) в продължение на 30 дни без приписваща токсичност. Artemisinins също имат някои противогъбични свойства срещу Pneumocystis carinii in vitro [100,101], въпреки че артеметер не е лечебен при две in vivo проучвания при имуносупресирани плъхове [102, 103] . Съществуват и няколко други модела на заболяването, като тези за ревматоиден артрит [104-106] , нефритен синдром [107] , панкреатит [108] и лупусен нефрит [109 , 110] , при които артемизинините са показали обещаващи резултати. В случай на лупусен нефрит, артемисинин се използва в продължение на три години в проучване при хора, с положителни ефекти върху болестното състояние [111] .Отидете на:
Заключителни бележки
Artemisinins са твърдо установени в комбинирани терапии [2,3] за лечение на резистентна към наркотици малария. Те се превръщат в анти-шистозомни агенти. Истинският им потенциал сега се крие в по-широките приложения за борба с болестите, особено при справянето с трудното предизвикателство, предизвикано от напредналите ракови заболявания, при които скъпото лечение осигурява в най-добрия случай постепенно нарастване на резултата. Въпросите относно режимите на дозиране, безопасността на продължителната употреба и възможните взаимодействия (положителни или отрицателни) със съществуващи терапии и токсични свойства, които биха могли да бъдат свързани с лечението на тумори, трябва да бъдат отговорени чрез подходящи клинични проучвания като част от спешната необходимост от изследване на лекарствата като например artesunate за онкологични показания. Отидете на:
Благодарности
Благодарим на Qinxue Hu за помощ при превеждането на много статии в китайски и Elvira Derbyshire за графики. SK благодарение на P. Folb, P. Kremsner, D. Kumar, Ajit Lalvani, Sir David Weatherall, Herwig Jensen и Tim Planche за изключителни съвети. RKH признава подкрепата на правителството на Фонда за облаги за високи научни постижения на Комитета за безвъзмездни средства за висши учебни заведения в Хонг Конг, проект № AoE P / 10-01-2-II и Съвета за безвъзмездни средства за университетите, грантове № HKUST 6493 / 06M и 600507. HMS е Wellcome Trust кариерен партньор (Grant No. 077441), спонсориран от SK. SK се финансира от Комисията на Европейските общности ANTIMAL Grant: 018834.Отидете на:
Бележки под линия
Обобщени таблици за ефектите на артемизинините върху видовете апикомплексани, други паразити и ракови заболявания са предоставени като допълнителни данни, които могат да бъдат намерени на doi: 10.1016 / jti.sps.2008.07.004 .Отидете на:
1. Zhang J.-F. Янг Чен Вечерни новини издателска компания; 2005. Подробен хронологичен запис на проект 523 и откриването и развитието на Qinghaosu (Artemisinin)2. Kremsner PG, Krishna S. Антималарийни комбинации. Lancet. 2004; 364 : 285-294. [ PubMed ]3. Бяла NJ антималариална лекарствена резистентност. J. Clin. Инвестирам. 2004; 113 : 1084-1092. [ PMC безплатна статия ] [ PubMed ]4. Haynes RK Разграждането с посредничеството на Fe2 + , свързването на PfATP6 и антималарийните активности на артемизона и други артемизинини: невъзможността на радикалите, центрирани с С, като биоактивни междинни продукти. Chem. Med. Chem. 2007; 2 : 1480-1497. [ PubMed ]5. Vennerstrom JL Идентифициране на кандидат за антималарично синтетично триоксоланно лекарство. Nature. 2004; 430 : 900-904. [ PubMed ]6. Gelb MH Откриване на наркотици за малария: едно много предизвикателно и навременно начинание. Curr. Opin. Chem. Biol. 2007; 11 : 440-445. [ PMC безплатна статия ] [ PubMed ]7. Golenser J. Текущи перспективи за механизма на действие на артемизинините. Int. J. Parasitol. 2006; 36 : 1427-1441. [ PubMed ]8. Jefford CW Новите разработки в синтетичните пероксидни лекарства, както имитира арттемизинин. Drug Discov. Today. 2007; 12 : 487-495. [ PubMed ]9. Кришна S. Преоценка на начина, по който артемисините действат в светлината на появяващите се доказателства за резистентност in vitro . Тенденции Mol. Med. 2006; 12 : 200-205. [ PMC безплатна статия ] [ PubMed ]10. Yuthavong Y. Malarial ( Plasmodium falciparum ) дихидрофолат редуктаза-тимидилат синтаза: структурна основа за антифолатна резистентност и развитие на ефективни инхибитори. Паразитология. 2005; 130 : 249-259. [ PubMed ]11. Yuvaniyama J. Забелязва антиполитична резистентност от маларийни DHFR-TS структури. Нат. Struct. Biol. 2003; 10 : 357-365. [ PubMed ]12. Питър У., Ричардс WHG 2-ро издание. Vol. 1. Springer-Verlag; 1984 (Handbook of Experimental Therapeutics Antimalarial Drugs).13. Питър У., Ричардс WHG 2nd edn. Vol. 2. Springer-Verlag; 1984 (Handbook of Experimental Therapeutics, Antimalarial Drugs).14. Хънт П. Ген, кодиращ деувиквитиниращ ензим, е мутирал в резистентни на артестенатни и хлорокини паразити на гризачи на малария. Mol. Microbiol. 2007; 65 : 27-40. [ PMC безплатна статия ] [ PubMed ]15. Афросо А. Маларийните паразити могат да развият устойчива резистентност към артемизинин, но нямат мутации в кандидат-гените atp6 (кодиращи саркоплазмичната и ендоплазмената ретикуларна Са2 + АТРаза), tctp, mdr1 и cg10 . Antimicrob. Агенти Chemother. 2006; 50 : 480-489. [ PMC безплатна статия ] [ PubMed ]16. Ferrer-Rodriguez I. Plasmodium yoelii : идентификация и частично охарактеризиране на MDR1 ген в резистентна към артемизинин линия. J. Parasitol. 2004; 90 : 152-160. [ PubMed ]17. Puri SK, Chandra R. Plasmodium vinckei : избор на щам, показващ стабилна устойчивост на артетера. Exp. Parasitol. 2006; 114 : 129-132. [ PubMed ]18. Walker DJ Механизми на резистентност към артемизинин в патогена на малария от гризачи Plasmodium yoelii. Antimicrob. Агенти Chemother. 2000; 44 : 344-347. [ PMC безплатна статия ] [ PubMed ]19. Bray PG Хинолини и артемизинин: химия, биология и история. Curr. Връх. Microbiol. Immunol. 2005; 295 : 3-38. [ PubMed ]20. Haynes RK Артемизинин антималариали не инхибират образуването на хемозини. Antimicrob. Агенти Chemother. 2003; 47 : 1175. [ PMC безплатна статия ] [ PubMed ]21. Eckstein-Ludwig U. Artemisinins са насочени към SERCA на Plasmodium falciparum . Nature. 2003; 424 : 957-961. [ PubMed ]22. Ellis DS Химиотерапията на маларията при гризачи XXXIX. Ултраструктурни промени след лечение с artemisinine на Plasmodium berghei инфекция при мишки, с наблюдения на локализацията на [ 3 Н] -dihydroartemisinine в P. фалципарум ин витро . Ан. Trop. Med. Parasitol. 1985; 79 : 367-374. [ PubMed ]23. Запаси PA Доказателство за общ механизъм за активиране, зависим от хелама, зависим от желязо, за полусинтетични и синтетични ендопероксидни антималарийни лекарства. Angew. Chem. Int. Ед. Engl. 2007; 46 : 6278-6283. [ PubMed ]24. Uhlemann AC Механизъм на антималариалното действие на синтетичния триоксолан RBX11160 (OZ277) Antimicrob. Агенти Chemother. 2007; 51 : 667-672. [ PMC безплатна статия ] [ PubMed ]25. Creek DJ Връзка между антималариалната активност и хематичното алкилиране на спиро- и диспиро-1,2,4-триоксалан антималарийни средства. Antimicrob. Агенти Chemother. 2008; 52 : 1291-1296. [ PMC безплатна статия ] [ PubMed ]26. Creek Kinetics на желязо-медиирано разграждане на артемизинин: ефект на състава на разтворителя и желязна сол. J. Pharm. Sci. 2005; 94 : 1820-1829. [ PubMed ]27. Charman SA Синтетични пероксидни антималариали от второ поколение. Am. J. Trop. Med. Hyg. 2007; 77 (Допълнение 5): 95.28. Kreidenweiss A. Антималарийна активност на синтетичен ендопероксид (RBx-11160 / OZ277) срещу изолати Plasmodium falciparum от Габон. Antimicrob. Агенти Chemother. 2006; 50 : 1535-1537. [ PMC безплатна статия ] [ PubMed ]29. Kannan R. Реакция на артемизинин с хемоглобин: последствия за антималариална активност. Biochem. J. 2005; 385 : 409-418. [ PMC безплатна статия ] [ PubMed ]30. ter Kuile F. Plasmodium falciparum: in vitro проучвания на фармакодинамичните свойства на лекарствата, използвани за лечение на тежка малария. Exp. Parasitol. 1993; 76 : 85-95. [ PubMed ]31. Desakorn V. Производство, зависимо от фаза, и освобождаване на богатия на хистидин протеин 2 от Plasmodium falciparum . Транс. R. Soc. Trop. Med. Hyg. 2005; 99 : 517-524. [ PubMed ]32. Charoenteeraboon J. Инактивиране на артемизинин чрез таласемични еритроцити. Biochem. Pharmacol. 2000; 59 : 1337-1344. [ PubMed ]33. Ittarat W. Ефекти на α-таласемията върху фармакокинетиката на антималариалния агент артесунат. Antimicrob. Агенти Chemother. 1998; 42 : 2332-2335. [ PMC безплатна статия ] [ PubMed ]34. Hutagalung R. Влияние на хемоглобин Е върху антималариалния ефект на производните на артемизинин. J. Infect. Dis. 2000; 181 : 1513-1516. [ PubMed ]35. Monti D. Дали хлорокинът действително действа чрез оксидативен стрес? FEBS Lett. 2002; 522 : 3-5. [ PubMed ]36. Parapini S. Доказателство, че желязото в маларийния паразит не е необходимо за антималариалните ефекти на артемизинин. FEBS Lett. 2004; 575 : 91-94. [ PubMed ]37. Jambou R. Резистентност на Field isolates на плазмодиум falciparum към in-vitro артеметерни и точкови мутации на PFATPase6 от тип SERCA. Lancet. 2005; 366 : 1960-1963. [ PubMed ]38. Legrand E. In vitro мониторинг на резистентността към лекарствения продукт на Plasmodium falciparum във Френска Гвиана: резюме на непрекъсната оценка от 1994 до 2005 г. Антимикроб. Агенти Chemother. 2008; 52 : 288-298. [ PMC безплатна статия ] [ PubMed ]39. Hayward R. pfmdr1 мутации, свързани с резистентност към хлорохина, понасят фитнес разходи в Plasmodium falciparum . Mol. Microbiol. 2005; 55 : 1285-1295. [ PubMed ]40. Walliker D. Фитнес на резистентни на наркотици паразити на малария. Акта Троп. 2005; 94 : 251-259. [ PubMed ]41. Cojean S. Устойчивост на дихидроартемизинин. Emerg. Заразяват. Dis. 2006; 12 : 1798-1799. [ PMC безплатна статия ] [ PubMed ]42. Legrand E. Устойчивост към дихидроартемизинин. Emerg. Заразяват. Dis. 2007; 13 : 808-809. [ PMC безплатна статия ] [ PubMed ]43. Toyoshima С. Кристална структура на калциевата помпа на саркоплазмичен ретикулум при 2.6 А резолюция. Nature. 2000; 405 : 647-655. [ PubMed ]44. Uhlemann AC Един аминокиселинен остатък може да определи чувствителността на SERCA към artemisinins. Нат. Struct. Mol. Biol. 2005; 12 : 628-629. [ PubMed ]45. Цена RN Резистентност към мефлоквин в Plasmodium falciparum и увеличен брой копия на ген на pfmdr1 . Lancet. 2004; 364 : 438-447. [ PMC безплатна статия ] [ PubMed ]46. Li W. Yeast модел разкрива двойни роли на митохондриите в действие на артемизинин. PLoS Genet. 2005; 1 : e36. [ PMC безплатна статия ] [ PubMed ]47. Alenquer М. Адаптивен отговор към антималарийния лекарствен артестенат в дрожди включва Pdr1p / Pdr3p-медиирано транскрипционно активиране на резистентните детерминанти TPO1 и PDR5 . FEMS Yeast Res. 2006; 6 : 1130-1139. [ PubMed ]48. Schmuck G. Невротоксичен начин на действие на артемизинин. Antimicrob. Агенти Chemother. 2002; 46 : 821-827. [ PMC безплатна статия ] [ PubMed ]49. Dondorp A. Artesunate срещу хинин за лечение на тежка фалципарна малария: рандомизирано проучване. Lancet. 2005; 366 : 717-725. [ PubMed ]50. Nealon C. Интрамускулна бионаличност и клинична ефикасност на артесунат при габонски деца с тежка малария. Antimicrob. Агенти Chemother. 2002; 46 : 3933-3939. [ PMC безплатна статия ] [ PubMed ]51. Woodrow CJ Artesunate срещу хинин за тежка falciparum малария: рандомизирано проучване. Lancet. 2006; 367 : 110-111. [ PubMed ]52. Gomes М. Rectal artemisinins for malaria: преглед на ефикасността и безопасността от отделните данни за пациентите в клинични проучвания. BMC Infect. Dis. 2008; 8 : 39. [ PMC безплатна статия ] [ PubMed ]53. Кришна С. Бионаличност и предварителна клинична ефикасност на интраректалния артесунат при деца от Гана с умерена малария. Antimicrob. Агенти Chemother. 2001; 45 : 509-516. [ PMC безплатна статия ] [ PubMed ]54. Campos, MS et al. Фатална токсичност при деца. J. Pediatr. Заразяват. Dis. (в пресата)55. Woodrow CJ Artemisinins. Postgrad. Med. J. 2005; 81 : 71-78. [ PMC безплатна статия ] [ PubMed ]56. Haynes RK Artemisone – високо активно антималариално лекарство от класа artemisinin. Angew. Chem. Int. Ед. Engl. 2006; 45 : 2082-2088. [ PubMed ]57. Nagamune K. Artemisinin индуцира калциево-зависима протеинова секреция в протозоичния паразит Toxoplasma gondii . Eukaryot. Cell. 2007; 6 : 2147-2156. [ PMC безплатна статия ] [ PubMed ]58. Nagamune K. Резистентните към артемизини мутанти на Toxoplasma gondii са променили калциевата хомеостаза. Antimicrob. Агенти Chemother. 2007; 51 : 3816-3823. [ PMC безплатна статия ] [ PubMed ]59. Флакон HJ, Gorenflot А. Химиотерапия срещу бабезиоза. Vet. Parasitol. 2006; 138 : 147-160. [ PubMed ]60. Utzinger J. Artemisinins за шистозомиаза и извън нея. Curr. Opin. Investig. Drugs. 2007; 8 : 105-116. [ PubMed ]61. Mishina YV Artemisinins инхибират Trypanosoma cruzi и Trypanosoma brucei rhodesiense in vitro растеж. Antimicrob. Агенти Chemother. 2007; 51 : 1852-1854. [ PMC безплатна статия ] [ PubMed ]62. Xiao SH Развитие на антишистозомните лекарства в Китай, като се обръща особено внимание на празиквантел и артезинини. Акта Троп. 2005; 96 : 153-167. [ PubMed ]63. Chen DJ Експериментални изследвания върху антишистозомната активност на qinghaosu. Zhong Hui Yi Xue Zha Zhi. 1980; 60 : 422-425.64. Efferth Т. Антималариумът artesunate е активен и срещу рака. Int. J. Oncol. 2001; 18 : 767-773. [ PubMed ]65. Келтър Г. Роля на трансфериновия рецептор и ABC транспортьорите ABCB6 и ABCB7 за резистентност и диференциация на туморни клетки към артеунтата. ЕДИНСТВЕНО. 2007; 2 : е798. [ PMC безплатна статия ] [ PubMed ]66. Li LN Artesunate атенюира растежа на човешки колоректален карцином и инхибира хиперактивния път на Wnt / β-катенин. Int. J. Cancer. 2007; 121 : 1360-1365. [ PubMed ]67. Zhou HJ Artesunate инхибира ангиогенезата и намалява експресията на съдовия ендотелен растежен фактор в клетките на К562 хронична миелоидна левкемия. Vascul. Pharmacol. 2007; 47 : 131-138. [ PubMed ]68. Wu ХН Дихидроартемизининът инхибира ангиогенезата, индуцирана от множествената миелома RPMI8226 клетки при хипоксични състояния чрез намаляване на експресията на съдовия ендотелен растежен фактор и подтискане на секрецията на съдовия ендотелен растежен фактор. Противоракови лекарства. 2006; 17 : 839-848. [ PubMed ]69. Li J., Zhou HJ Dihydroartemisinin инхибира експресията на съдовия ендотелен растежен фактор в К562 клетки. Яо Xue Xue Бао. 2005; 40 : 1041-1045. [ PubMed ]70. Anfosso L. Microarray експресионните профили на свързаните с ангиогенезата гени предсказват отговора на туморни клетки към артемизинини. Pharmacogenomics J. 2006; 6 : 269-278. [ PubMed ]71. Longo M. Ефекти на антималариалното лекарство дихидроартемизинин (DHA) върху ембриони на плъхове in vitro . Reprod. Toxicol. 2006; 21 : 83-93. [ PubMed ]72. Dell’Eva R. Инхибиране на ангиогенезата in vivo и растежа на ксенографски тумори на саркома на Kaposi от анти-маларийния артестенат. Biochem. Pharmacol. 2004; 68 : 2359-2366. [ PubMed ]73. Huan-huan C. Artesunate намалява неоваскуларизацията на пилешки хориоалантоични мембрани и проявява антиангиогенна и апоптотична активност върху човешки микроваскуларни кожни ендотелни клетки. Cancer Lett. 2004; 211 : 163-173. [ PubMed ]74. Chen HH Антималариалният дихидроартемизинин също инхибира ангиогенезата. Cancer Chemother. Pharmacol. 2004; 53 : 423-432. [ PubMed ]75. Chen HH Инхибиторни ефекти на артесуната върху ангиогенезата и върху експресията на съдовия ендотелен растежен фактор и VEGF рецептора KDR / flk-1. Фармакология. 2004; 71 : 1-9. [ PubMed ]76. Wartenberg М. Антималарийното средство artemisinin упражнява антиангиогенен ефект в ембрионалните органи, получени от миши ембрионални стволови клетки. Lab. Инвестирам. 2003; 83 : 1647-1655. [ PubMed ]77. Chen HH Инхибиране на растеж на човешка ракова клетъчна линия и ангиогенеза на ендотелиални клетки от човешка пъпна връв от производни на артемизинин in vitro . Pharmacol. Res. 2003; 48 : 231-236. [ PubMed ]78. Wu GD Апоптоза на човешки ендотелни клетки от пъпна вена, индуцирана от артещунат. Vascul. Pharmacol. 2004; 41 : 205-212. [ PubMed ]79. Efferth Т., Oesch F. Оксидативен стрес отговор на туморни клетки: основаващо се на микрочипове сравнение между артемизинини и антрациклини. Biochem. Pharmacol. 2004; 68 : 3-10. [ PubMed ]80. Efferth T. Ролята на антиоксидантните гени за активността на артестенатните срещу туморните клетки. Int. J. Oncol. 2003; 23 : 1231-1235. [ PubMed ]81. Singh NP, Lai HC Синергична цитотоксичност на артемизинин и натриев бутират върху човешки ракови клетки. Anticancer Res. 2005; 25 : 4325-4331. [ PubMed ]82. Kim SJ Dihydroartemisinin повишава радиоактивността на клетките на глиома на човек in vitro . J. Cancer Res. Clin. Oncol. 2006; 132 : 129-135. [ PubMed ]83. Efferth Т. Подобряване на цитотоксичността на артемизинини спрямо ракови клетки чрез желязо. Безплатно Radic. Biol. Med. 2004; 37 : 998-1009. [ PubMed ]84. Singh NP, Lai HC Artemisinin индуцира апоптоза в човешките ракови клетки. Anticancer Res. 2004; 24 : 2277-2280. [ PubMed ]85. Sadava D. Трансгрин преодолява лекарствената резистентност към артемизинин в човешки клетки на белодробна белодробна карцинома. Cancer Lett. 2002; 179 : 151-156. [ PubMed ]86. Singh NP, Lai H. Селективна токсичност на дихидроартемизинин и холотрансферин към човешки ракови клетки на гърдата. Life Sci. 2001; 70 : 49-56. [ PubMed ]87. Lai H., Singh NP Селективна цитотоксичност на раковите клетки от излагане на дихидроартемизинин и холотрансферин. Cancer Lett. 1995; 91 : 41-46. [ PubMed ]88. Berger TG Artesunate при лечението на метастатичен увеален меланом – първи опит. Oncol. Rep2005; 14 : 1599-1603. [ PubMed ]89. Singh NP, Panwar VK Доклад за хипофизния макроаденом, лекуван с артеметер. Integr. Рак Тер. 2006; 5 : 391-394. [ PubMed ]90. Zhang ZY Artesunate комбинирани с винорелбин плюс цисплатин при лечение на напреднал недребноклетъчен белодробен рак: рандомизирано контролирано проучване. Zhong Xi Yi Jie Той Xue Бао. 2008; 6 : 134-138. [ PubMed ]91. Paeshuyse J. Hemin потенцира анти-хепатит С вирусната активност на антималарийното лекарство artemisinin. Biochem. Biophys. Res. Commun. 2006; 348 : 139-144. [ PubMed ]92. Romero MR Ефект на artemisinin / artesunate като инхибитори на продукцията на вируса на хепатит В в “ in vitro “ репликативна система. Antiviral Res. 2005; 68 : 75-83. [ PubMed ]93. Efferth Т. Антивирусна активност на артестенат спрямо див тип, рекомбинантни и ганцикловир-резистентни човешки цитомегаловируси. J. Mol. Med. 2002; 80 : 233-242. [ PubMed ]94. Kaptein SJ Анти-маларийният медикамент artesunate инхибира репликацията на цитомегаловирус in vitro и in vivo . Antiviral Res. 2006; 69 : 60-69. [ PubMed ]95. Naesens L. Антивирусна активност на различни класове широко действащи средства и естествени съединения при HHV-6-инфектирани лимфобласти. J. Clin. Virol. 2006; 37 (Допълнение 1): S69-S75. [ PubMed ]96. Efferth Т. Дейност на лекарства от традиционната китайска медицина към чувствителни и MDR1- или MRP1-свръхекспресиращи мултирезистентни човешки CCRF-CEM левкемични клетки. Кръвни клетки Mol. Dis. 2002; 28 : 160-168. [ PubMed ]97. Qian RS Имунологичен и антивирусен ефект на qinghaosu. J. Tradit. Брадичка. Med. 1982; 2 : 271-276. [ PubMed ]98. Romero MR Антивирусен ефект на artemisinin от Artemisia annua срещу модел, член на фамилията Flaviviridae, вирус на говежди вирусна диария (BVDV) Planta Med. 2006; 72 : 1169-1174. [ PubMed ]99. Shapira MY Artesunate като мощен антивирусен агент при пациент с късна лекарствена резистентна цитомегаловирусна инфекция след трансплантация на хемопоетични стволови клетки. Clin. Заразяват. Dis. 2008; 46 : 1455-1457. [ PubMed ]100. Merali S., Meshnick SR податливост на Pneumocystis carinii към artemisinin in vitro . Antimicrob. Агенти Chemother. 1991; 35 : 1225-1227. [ PMC безплатна статия ] [ PubMed ]101. Ni X., Chen Y. In vitro изследване на ефекта на анти-пневмоцистит карнини на производни на артемицин. Zhonghua Jie Той Той Ху Xi Za Zhi. 2001; 24 : 164-167. [ PubMed ]102. Brun-Pascaud M. Липса на активност на артеметер за профилактика и лечение на инфекции с Toxoplasma gondii и Pneumocystis carinii при плъхове. Паразит. 1996; 3 : 187-189. [ PubMed ]103. Chen YT Експериментално изследване на Artemether при лечение на Pneumocystis carinii при имуносупресирани плъхове. Брадичка. Med. J. (Engl.) 1994; 107 : 673-677. [ PubMed ]104. Xu H. Анти-маларийният агент артесунат инхибира индуцираната от TNF-a продукция на провъзпалителни цитокини чрез инхибиране на NF-кВ и PI3 киназа / Akt сигнален път в човешки ревматоиден артрит фибо-бласт-подобни синовиоцити. Ревматология (Оксфорд) 2007; 46 : 920-926. [ PubMed ]105. Mirshafiey A. Проектиране на нова линия при лечение на експериментален ревматоиден артрит от артестант. Immunopharmacol. Immunotoxicol. 2006; 28 : 397-410. [ PubMed ]106. Cuzzocrea S. Artemether: нова терапевтична стратегия при експериментален ревматоиден артрит. Immunopharmacol. Immunotoxicol. 2005; 27 : 615-630. [ PubMed ]107. Razavi A. Лечение на експериментален нефротичен синдром с артеунтата. Int. J. Toxicol. 2007; 26 : 373-380. [ PubMed ]108. Zhao M. Индукция на апоптоза от артемизинин, облекчаваща тежестта на възпалението при остър панкреатит, индуциран от карелеин. Свят J. Gastroenterol. 2007; 13 : 5612-5617. [ PMC безплатна статия ] [ PubMed ]109. Li WD Dihydroarteannuin подобрява симптома на лупус на BXSB мишки чрез инхибиране на продукцията на TNF-a и блокиране на NF-кБ транслокацията на сигналната пътека. Int. Immunopharmacol. 2006; 6 : 1243-1250. [ PubMed ]110. Dong YJ Ефект на дихидро-qinghaosu върху производството на авто-антитела. TNFa секреция и патологична промяна на лупусния нефрит при BXSB мишки. Zhongguo Zhong Xi Yi Jie Той За Zhi. 2003; 23 : 676-679. [ PubMed ]111. Lu L. Изследване на ефекта от Cordyceps sinensis и artemisinin за предотвратяване на рецидиви на лупусния нефрит. Zhongguo Zhong Xi Yi Jie Той За Zhi. 2002; 22 : 169-171. [ PubMed ]112. Li Q. Токсикокинетика и хидролиза на артелинат и артестенат при плъхове, инфектирани с малария. Int. J. Toxicol. 2005; 24 : 241-250. [ PubMed ]
Снимка: Червените кръвни клетки са заразени с маларийни паразити в рода Plasmodium. Credit: Kateryna Kon / Shutterstock.comBy Jeanna Bryner, Live Science Managing Editor
Нобеловата награда за физиология и медицина от 2015 г. е присъдена на трио от учени за открития, които доведоха до нови лечения за някои от най-опустошителните паразитни болести, съобщи фондация „Нобелова“ тази сутрин (5 октомври).
Половината от Нобеловата награда бе присъдена съвместно на Уилям К. Кембъл и Сатоши Омура за откриване на ново лечение за инфекции, причинени от паразити на кръгли червеи. Другата половина отиде в Youyou Tu за откриване на наркотици за борба с маларията, болестта, предавана от комари, която отнема около 450 000 живота годишно в световен мащаб, според Нобеловата фондация.
„Тези две открития са предоставили на човечеството мощни нови средства за борба с тези инвалидизиращи заболявания, които засягат стотици милиони хора годишно. Последиците за подобряване на човешкото здраве и намаленото страдание са неизмерими“, заявиха представители на Нобеловата фондация в изявление . [ 7 Революционни Нобелови награди в медицината ]
Кембъл и Омура са открили лекарствения авермектин, чиито производни са спомогнали за лечение на речна слепота и лимфатична филариаза. Риборната слепота причинява възпаление на роговицата и евентуална слепота, а лимфната филариаза причинява хронично подуване и може да доведе до елефантиаза (крайно подуване на ръцете и краката) и скротален хидроцеле (подуване на скротума).
Омура, микробиолог и професор в университета „Китасато“ в Япония, направи първата крачка към откриването на печелившия Нобелов лауреат. Той знае, че много бактериални видове от рода Streptomyces показват антибактериални свойства. Òmura успя да изолира нови щамове Streptomyces от почвата и успешно да ги култивира в лабораторията. От тези Омура избра около 50 от най-обещаващите щамове за по-нататъшно тестване.
Тогава Кембъл, който в момента е научен сътрудник в Университета Дрю в Медисън, Ню Джърси, показа, че компонент в една от тези култури е ефективен при борбата с паразити в домашни и селскостопански животни. Този компонент се пречиства в лекарствения авермектин, който след това се модифицира в съединението Ivermectin. Лекарството Ivermectin по-късно е установено, че ефективно убива паразитни ларви.
Търсенето от типа на антималарични агенти започна в края на 60-те години на миналия век, във време, когато болестта се увеличаваше; традиционните методи за лечение на малария – хлорохин и хинин – стават все по-малко ефективни. Ту, главен професор в Китайската академия за традиционна китайска медицина, се обърна към билковото лекарство и започна да преглежда тези билки при животни, заразени с малария. Тя откри един екстракт от растението Artemisia annua, който изглеждаше обещаващ.
След извличането на активния компонент от растението, Ту показва, че е ефективен срещу маларийния паразит при животни и хора. Това съединение сега се нарича Aртемисинин.
Камбъл и Омура ще споделят половината от тази година с Нобелова награда от 8 милиона шведски крони (около 960 000 долара), а Ту ще получи другата половина от парите.
Този сайт използва „бисквитки“, за да работи оптимално за Вас. Чрез посещението си Вие приемате използването на „бисквитки“.